El flagelo sigue exhibiendo complejidad irreducible — Michael Behe no ha sido refutado
Jonathan M
Los que han ido siguiendo el debate entre la tesis del Diseño Inteligente por una parte y el darwinismo por otra durante un cierto tiempo estarán bien familiarizados con la réplica estándar de los darwinistas respecto del argumento de Behe de la complejidad irreducible que se exhibe en el flagelo bacteriano. Parece haber unanimidad entre los teóricos del darwinismo de que el argumento de la complejidad irreducible en el flagelo bacteriano ha sido refutado, y que los proponentes del Diseño Inteligente estamos constantemente cambiando las reglas, hundiendo la cabeza en la arena y en general agarrándonos a un clavo ardiendo. Por ejemplo, un participante en Facebook observó recientemente:
Mi principal queja acerca de los proponentes del Diseño Inteligente es que parece que nunca cejan. ¿Cuántas veces se le debe decir a alguien que está equivocado hasta que lo admita? ¿Cuántas veces se tiene que refutar el Diseño Inteligente en los medios sometidos a revisión por pares antes que se abandone como una causa perdida? La historia de la complejidad irreducible del flagelo bacteriano está total y completamente muerta. Es un error. Abandonadla.
El caso es que recientemente saqué a colación el flagelo como un caso documentado de complejidad irreducible en un Coloquio sobre ciencia y fe, y recibí respuestas de un tenor muy parecido.
Pero, ¿es cierta esta afirmación? De verdad ha sido refutado este argumento por sus críticos? Hará un año que leí Por qué fracasa el Diseño Inteligente — Una crítica científica del Nuevo Creacionismo (editado por Matt Young y Taner Edis). El autor del Capítulo 5 de dicha obra es Ian Musgrave y se titula «La evolución del flagelo bacteriano». Escribiendo en respuesta a Michael Behe y William Dembski , Musgrave intenta disipar la noción de la complejidad irreducible de una vez por todas. Recuerdo que al leer este capítulo no me sentí en absoluto convencido por sus argumentos. En la página 82 del libro, Musgrave nos ofrece el siguiente argumento:
Aquí tenemos un posible escenario para la evolución del flagelo eubacteriano: primero surgió un sistema de secreción, basado alrededor del vástago SMC y del complejo conformador de poros, que fue el antecesor común del sistema de secreción tipo III y del sistema flagelar. La asociación de una bomba iónica (que posteriormente llegó a ser la proteína motora) a esta estructura mejoró la secreción. Incluso actualmente, las proteínas motoras, parte de una familia de proteínas impulsoras de secreción, pueden disociarse y reasociarse libremene en esta etapa, como sucede en algunos sistemas de motilidad por deslizamiento. El filamento protoflagelar surgió a continuación como parte de la estructura de secreción de proteínas (comparar el pilus de la Pseudomonas, los apéndices filamentosos de la Salmonella, y las estructuras filamentosas de la E. coli). La motilidad de deslizamiento-contracción surgió en esta etapa o posteriormente, y luego fue afinada a una motilidad natatoria. La regulación y la conmutación pueden añadirse después, porque hay modernas eubacterias que carecen de estos atributos pero que funcionan bien en sus medios. (Shah y Sockett 1995). En cada etapa hay un beneficio a los cambios en la estructura.
De hecho, Mark Pallen y Nick Matzke formulan un argumento muy parecido en su artículo deNature Reviews de 20061 (un artículo que fue citado por un miembro de la audiencia durante la reciente gira de Behe en el Reino Unido). Ken Miller es también conocido por realizar afirmaciones parecidas acerca de una evolución del flagelo a partir del Sistema de Secreción de Tipo III basándose principalmente en consideraciones de homologías de secuencias proteínicas.
Entonces, ¿se consigue con estos alegatos refutar de una vez por todas el inconveniente argumento del diseño inteligente? Bien, la verdad es que no, en absoluto. De hecho, propongo que los argumentos de todos los autores citados trivializan de forma fundamental diversas cuestiones de gran importancia.
Primero, y sobre todo, trivializa la pasmosa complejidad y sofisticación del sistema flagelar —tanto su aparato de ensamblaje como su vanguardista diseño en sí mismo. En realidad, el proceso por el que se realiza el proceso de autoensamblaje del flagelo bacteriano dentro de la célula es tan sofisticado que he encontrado siempre que cuesta exponerlo de una manera accesible a los legos en la materia. Sus conceptos fundamentales son notoriamente difíciles de captar por parte de los que no están acostumbrados a pensar acerca del sistema, o por parte de los que lo abordan por primera vez. Pero, al mismo tiempo, la base mecanística del ensamblaje del flagelo es tan pasmosamente elegante y fascinante que la absoluta genialidad del diseño técnico del motor flagelar —y desde luego la magnitud del desafío que planeta al darwinismo— no pueden apreciarse idóneamente sin como mínimo un conocimiento elemental de las operaciones subyacentes. Echemos un vistazo.
El autoensamblaje del aparato flagelar
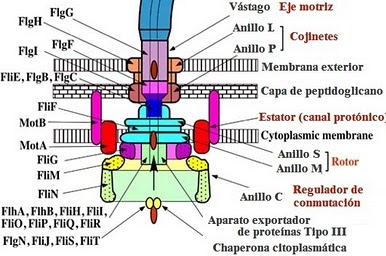
La síntesis del flagelo bacteriano demanda la expresión coordinada de más de 60 productos genéticos. Su biosíntesis en el interior de la célula es orquestada por genes organizados en una cascada sumamente ordenada en la que la expresión de un gen a un nivel determinado exige la previa expresión de otro gen a un nivel más elevado. El organismo paradigmático, o modelo, para el ensamblaje del flagelo es la Salmonella, una bacteria de la familiaEnterobacteriaceae. Mis consideraciones así se referirán principalmente a la Salmonella, excepto si se indica lo contrario.
El sistema flagelar en la Salmonella tiene tres clases de promotores (los promotores son una especie de conmutador molecular que puede iniciar la expresión de los genes cuando son reconocidos por la ARN-polimerasa y una proteína especializada asociada llamada «factor sigma»). Estas tres clases de promotores son designados de manera bien simple como «Clase I», «Clase II» y «Clase III». Esta transcripción secuencial está vinculada con el proceso del ensamblaje del flagelo. La Clase I contiene sólo dos genes en un operón (llamado FlhD y FlhC). La Clase II consiste de 35 genes incorporados en ocho operones (incluyendo genes involucrados en el ensamblaje del Cuerpo Basal del Codo y de otros componentes del flagelo, así como del aparato de exportación y dos genes reguladores llamados «FliA» y «FlgM»). Estos genes involucrados en la síntesis del filamento están controlados por los promotores de la Clase III.
El promotor de Clase I activa la expresión de un regulador maestro (privativo de lasEnterobacteriaceae de las que la Salmonella es un miembro) llamado «FldH4C2» (¡no es necesario memorizar el nombre!). Este regulador entérico maestro activa luego los promotores de Clase II en asociación con un factor sigma, σ70 (recordemos que un factor sigma es un tipo de proteína que habilita la unión específica de ARN-polimerasa a promotores génicos). Los promotores de Clase II son luego responsables de la expresión génica de las subunidades del cuerpo basal del codo y de sus reguladores, incluyendo otro factor sigma, el σ28 (que está codificado por un gen llamado FliA) y su factor anti-sigma, el FlgM (los factores anti-sigma, como lo sugiere su nombre, se unen a los factores sigma para inhibir su actividad transcripcional). El factor sigma σ28 es necesario para activar los promotores de Clase III. Pero aquí nos encontramos con un problema potencial. No tiene ningún sentido comenzar a expresar los monómeros de la flagelina antes de completar la construcción del Cuerpo Basal del Codo. Así, para inhibir el σ28, el factor anti-sigma (FlgM) antes mencionado inhibe su actividad y le impide interaccionar con el complejo holoenzimático ARN-polimerasa. Cuando se ha completado la construcción del Cuerpo Basal del Codo, se segrega el factor anti-sigma FlgM a través de las estructuras flagelares que se producen por la expresión de los genes de Clase II del cuerpo basal del codo. Los promotores de Clase III (responsables de la expresión de los monómeros de la flagelina, del sistema de quimiotaxis y de los generadores de fuerza motriz) son luego por fin activados por el factor σ28, y se puede proceder a completar el flagelo.
Pero hay mucho más. El sistema de exportación del flagelo (es decir, el medio por el que la FlgM es extraído desde la célula) tiene dos estados sustrato-específicos: sustratos tipo vástago/codo, y sustratos tipo filamento. Durante el proceso del ensamblaje del flagelo, este conmutador de especificidad de sustrato tiene que pasar del primero de estos dos estados al otro. Las proteínas que forman parte del codo y del vástago tienen que ser exportadas antes que las que conforman el filamento. Pero, ¿cómo tiene lugar esta conmutación en la especificidad del sustrato?
El participante clave en este proceso es una proteína de la membrana llamada FlhB. Hay también una proteína flagelar que posee la longitud del codo y que es responsable de cerciorarse de que la longitud del codo es la correcta (alrededor de 55nm) llamada FliK. Esta misma proteína es también la responsable de iniciar la conmutación de exportación de la especificidad del sustrato. Sucede que sin FliK se pierden totalmente tanto la capacidad de conmutar y exportar filamento así como la de controlar la longitud del codo. FliK tiene dos dominios clave, esto es, los dominios N-terminal y C-terminal. Durante el ensamblaje del codo, FliKN funciona como un sensor y transmisor molecular de información sobre la longitud del codo. Cuando el codo alcanza la longitud adecuada, la información se transmite a FliKC y a FliKCT, lo que resulta en un cambio de conformación que a su vez resulta en que FliKCT se une a FlhBC. Esto, a su vez, resulta en un cambio de conformación en FlhBC. Esto causa la conmutación de especificidad del sustrato.
El ensamblaje del flagelo comienza en la membrana citoplasmática, progresa a través del espacio periplasmático y se extiende finalmente al exterior de la célula. Fundamentalmente, el flagelo se compone de dos partes principales: el sistema de secreción y la estructura axial. Los principales componentes de la estructura axial son FlgG para el vástago, FlgE para el codo, y FliC para el filamento. Todos estos se ensamblan con la ayuda de una proteína tapón (FlgJ, FlgD y FliD respectivamente). De éstas, sólo FliD permanece en el extremo del filamento del producto acabado. Otros componentes de la estructura axial (llamados FlgB, FlgC y FlgF) conectan el vástago y el complejo del anillo MS. El codo y el filamento están conectados por FlgK y FlgL.
Cuando el anillo C y el vástago C se unen al anillo M en su superficie citoplasmática, el complejo del anillo MS —que es el fundamento estructural del aparato— puede comenzar a segregar proteínas flagelares.
La estructura del vástago se construye a través de la capa de peptidoglicano. Pero el crecimiento no puede proseguir sin ayuda más allá de la barrera física que presenta la membrana exterior. De modo que el complejo anular exterior acabado de mencionar corta un orificio en la membrana, de modo que el codo puede crecer debajo del andamiaje de la FlgD hasta que alcanza la longitud crítica de 55nm. Entonces los sustratos que están siendo segregados pueden pasar desde el modo vástago-codo al modo flagelina, la FlgD puede ser sustituida por proteínas asociadas con el codo, y el filamento sigue creciendo. Sin la presencia de la proteína tapón FliD, estos monómeros de flagelina se pierden. Así, esta proteína tapón es esencial para que el proceso pueda tener lugar.
Por qué no es posible una evolución del flagelo a partir del T3SS
Sería de pensar que la descripción acabada de dar del sistema y de su autoensamblaje sería más que suficiente para dejar a descubierto cómo los juegos de mano de Kenneth Miller et al. se limitan a trivializar la cuestión hasta el extremo. Pero las cosas se ponen peor todavía para la narrativa darwinista. ¿Por qué exactamente está la biosíntesis del flagelo tan extremadamente regulada y coordinada? No sólo las demandas energéticas hacen del flagelo un sistema muy caro de funcionar, sino que una expresión fuera de tiempo de las proteínas del flagelo pueden inducir una enérgica respuesta inmune en el sistema huésped, algo que ninguna bacteria quiere experimentar.
¿Cuál es el sentido de esto desde el punto de vista evolucionista? Bien, los monómeros de flagelina son unos inductores bastante potentes de la citocina. Si resulta que eres un organismo Yersinia poseedor de un Sistema de Secreción Tipo III, lo último que quieres es exhibir estos péptidos de flagelina a los macrofagos. Estos podrían cancelar de manera significativa la estrategia antiinflamatoria de la Yersinia.
Conclusión
En realidad, la descripción que he presentado más arriba no hace más que arañar la superficie de este espectacular artículo nanotecnológico2. Para abreviar, ni tan siquiera he considerado el extraordinario proceso de la quimiotaxis, los circuitos de transducción de señales de dos componentes, la conmutación giratoria y la fuerza motriz protónica mediante la que se alimenta al flagelo. Pero la realidad fundamental es que la teoría darwinista —tal como se comprende de forma clásica— no se ha aproximado siquiera a explicar el origen de este extraordinariamente complejo y sofisticado motor. Así como las «explicaciones» darwinistas del ojo pueden parecer de entrada convincentes a los desconocedores de la complejísima ingeniería de diseño de la base bioquímica y molecular de la visión, también las «explicaciones» evolucionistas del flagelo quedan vacías de toda capacidad persuasiva cuando uno considera los detalles moleculares del sistema. Cuando se unen los anteriores detalles con demostraciones de la absoluta impotencia de la tesis neodarwinista para producir novedosos pliegues proteínicos y nuevos lugares de unión proteína-proteína, ¿se puede realmente mantener que este sistema se puede conseguir en virtud de ligeras modificaciones sucesivas, pasito a pasito? Dado que el principal argumento darwinista reside en su pretendida eficacia en desvirtuar la abrumadora apariencia de designio, ¿no es razonable que la demostrable impotencia de dicho argumento darwinista devuelve el postulado del designio a la mesa como una proposición científica viable y respetable?
Douglas Axe, del Instituto Biológico, exponía en un reciente artículo en la revista Bio-complexity3 que el modelo de duplicación y movilización genética sólo funciona si se precisa de muy pocos cambios para adquirir una utilidad novedosa seleccionable o neo-funcionalización. Si un gen duplicado es neutral (en términos de su coste para el organismo), entonces la máxima cantidad de mutaciones que puede necesitar una innovación novedosa en una población bacteriana es de hasta seis. Si el gen duplicado tiene un coste ligeramente negativo relativo a la aptitud, la cantidad máxima cae a dos o menos (aparte de la duplicación misma).
Es patente que el flagelo bacteriano sigue siendo un reto tan poderoso para el darwinismo —y quizá mucho más que cuando Behe publicó su libro La Caja Negra de Darwin en 1996.
- Nature – From The Origin of Species to the origin of bacterial flagella – October 2006
- FEMS Microbiology reviews – Regulation cascade of Fagellar expression in Gram-negative bacteria – First published online 16 June 2003
- BIO-Complexity, Vol 2010 – The Limits of Complex Adaptation: An Analysis Based on a Simple Model of Structured Bacterial Populations
Evolution News – Michael Behe Hasn’t Been Refuted on the Flagellum – March 15, 2011
Extr de http://www.creacionismo.net/genesis/Art%C3%ADculo/el-flagelo-sigue-exhibiendo-complejidad-irreducible-%E2%80%94-michael-behe-no-ha-sido-refutado
Con mi mayor respeto. Como mi coeficiente intelectual creo que es mediano, intuyo que el artículo podría ser interesante a nivel general si se bajara un cambio o un peldaño en el desarrollo del tema; de lo contrario creería que es tiempo y espacio perdido su inclusión, excepto para un neófito.
Básicamente se trata de encontrar, y resaltar, puntos débiles a la teoría de la evolución para así » justificar » el creacionismo.
La teoría de la evolución tiene puntos débiles, por éso sigue siendo una teoría. Si no tuviese aspectos aún inexplicados sería una Ley.
Pero demostrar ( si acaso se logra ) que algo no es azul, no lo convierte necesaria e irremediablemente en verde.
Que haya aspectos determinados y concretos en los que la teoría de la evolución no aporta una respuesta sólida, no significa que sea cierto el designio divino.
Probablemente la teoría de la evolución está incompleta o incluso puede ser errónea en cuanto a su funcionamiento ( así lo pienso yo ) que no en tanto a su naturaleza. Pero para afirmar el creacionismo es necesario dar respuestas a muchas preguntas que éste plantea. Y éso no se hace.
Se echa de menos que alguien aplique el mismo rigor y puntillismo a las tesis creacionistas. En vez de tratar de desmontar la teoría aceptada, lo que deberían hacer es argumentar la propuesta alternativa, pero…………….es completamente imposible.
¿Encontrar lugares oscuros en la teoría de la evolución nos sirve para abrazar el creacionismo de manera acrítica ? A mí no.
Gracias José. Lo planteaste magníficamente.